Verde Casino
Intră în lumea captivantă a jocurilor de noroc cu Verde Casino, o destinație online ce redefinește experiența cazinourilor pentru jucătorii din România. Lansată în 2022, platforma noastră nu doar aduce prospetime în industrie, ci și se angajează să ofere o alternativă excepțională în ceea ce privește diversitatea jocurilor, câștigurile generoase și o gamă impresionantă de bonusuri și promoții. Deși suntem o prezență relativ nouă, ne-am dedicat să aducem laolaltă cele mai moderne facilități, inclusiv procese rapide de înregistrare, opțiuni de bonus captivante, un program de loialitate bogat în recompense cash, rotiri gratuite și cashback generos în mii de Ron. Cu jocuri oferite de producători renumiți în industria sloturilor și a jocurilor de masă, alături de o experiență de cazino live la superlativ, casino Verde își propune să transforme fiecare pariu într-un moment memorabil. Bine ai venit într-o comunitate unde distracția și câștigurile se întâlnesc într-un mod excepțional!
Verde casino — recenzie
Descoperă o experiență de cazino aproape de natură cu Verde Casino, o destinație inovatoare ce aduce o temă incredibil de proaspătă în lumea jocurilor de noroc. Operat de către compania Brivio Limited, înregistrată în Cipru, acest operator este licențiat de Invicta Networks sub numărul de licență 8048/JAZ2012-009, garantând astfel jocuri corecte și de încredere pentru pasionații de cazino. Lansat în 2022 pentru piața internațională a jocurilor de noroc online, casino Verde se remarcă prin parteneriatele cu unele dintre cei mai buni furnizori de jocuri de noroc, aducând o varietate captivantă de opțiuni în secțiunile sale diverse de jocuri. Cei care aleg să exploreze această platformă vor găsi o gamă extinsă de jocuri în meniul Lobby, beneficiind de acces la cele mai noi promoții și având posibilitatea de a descoperi constant jocuri noi și palpitante. Cu o prezență activă în mai multe limbi, inclusiv engleză și română, Verde casino RO oferă o experiență personalizată pentru jucătorii din diverse colțuri ale lumii. Fiind dedicat transparenței și divertismentului responsabil, acest casino aduce o doză proaspătă și captivantă în universul jocurilor de noroc online.
| 🔥 Licenta | 8048/JAZ2012-009 |
| 🎯 Bonus | 4800 RON și 220 de rotiri gratuite |
| 💎 Bonus Fara depunere | Da |
| 💸 Live Chat | Nu |
| 💵 Verde Casino | Joacă aici! |
| 🎲 Suma min la depunere | 30 RON |
| 💰 Numărul total de jocuri | 1001+ |
| 🧭 Pachet bonus | Ia Bonusul! |
| ⭐ Opțiuni de depunere | Visa, MasterCard |
| 🤑 Furnizor de software | EGT, Evolution |
Avantajele și dezavantajele Verde Casino
| Avantaje | Defecte |
| Operare sub licență | Lipsa unei aplicații pentru iPhone |
| Bonus generos de bun venit | Variație limitată a promoțiilor |
| Program de fidelizare | |
| Diverse sisteme de plată | |
| Jocuri de cazinou live |
Verde Casino înregistrare cont
Pentru a profita de toate beneficiile oferite de Verde Casino, inclusiv securitate avansată și oferte exclusive, trebuie să te înregistrezi ca membru. Iată pașii simpli pe care trebuie să îi urmezi pentru a-ți crea contul de jucător:
- Pasul 1: Accesează site-ul Verde casino RO făcând click pe oricare dintre ofertele prezentate pe platformă.
- Pasul 2: Completează formularul de înregistrare cu informațiile tale personale. Asigură-te că oferi date corecte și valide.
- Pasul 3: Crează contul și validează-l. Verifică-ți adresa de email sau numărul de telefon pentru a confirma înregistrarea.
- Pasul 4: După validare, revendică bonusul de bun venit oferit și începe să explorezi jocurile oferite de Verde Casino.
Pentru o înregistrare rapidă, poți opta să te conectezi folosind un cont de social media precum Facebook, Twitter, TikTok sau Google.
Atenție: Este esențial să respecți regulile de utilizare și să furnizezi informații corecte pentru a asigura o experiență de joc sigură și transparentă.
Autentificare în cont
Pentru a accesa contul tău Verde Casino, navighează către butonul “Autentificare” situat în partea superioară a paginii principale. Completează cu datele tale de autentificare, inclusiv numele de utilizator sau adresa de email și parola asociată. Apasă butonul “AUTENTIFICARE” pentru a te conecta rapid și ușor la experiența ta de joc.
Cele mai bune bonusuri și oferte Verde Casino

În lumea cazinourilor online, bonusurile reprezintă o modalitate atrăgătoare prin care jucătorii noi și cei fideli sunt răsplătiți. Verde Casino nu face excepție și impresionează cu o gamă diversificată de oferte atractive. Iată câteva exemple de bonusuri pe care le poți întâlni în această platformă:
- Cashback Atractiv:
- Interval cashback între 3% și 12%
- Promoții speciale din rețeaua Verde Casino
- Oferte Premium pentru VIP-uri:
- Bonusuri fără depunere
- Cashback exclusiv
- Oferte personalizate
- Bonusuri Sezoniere:
- Bonusuri tematice pentru sărbători (Crăciun, Paște, etc.)
- Roata Norocului:
- Oportunitatea de a câștiga premii instant
- Drops & Wins Promo:
- Promoții cu premii și câștiguri
- Turnee Săptămânale și Lunare:
- Competiții periodice cu premii atractive
- Jackpoturi și Bonusuri Fără Depunere Speciale:
- Șanse de câștig la jackpoturi
- Oferte speciale fără a depune sume
- Ofertă Specială cu Cod Promoțional pentru Rotiri Gratuite:
- Rotiri gratuite cu coduri promoționale exclusive
Este esențial să fii conștient că majoritatea acestor bonusuri sunt supuse unor termeni și condiții. Acestea includ cerințe de rulaj, condiții de pariere, cashout maxim, limite de pariere, selecții specifice de jocuri, și altele. Prin urmare, este recomandat să citești cu atenție regulile asociate fiecărei oferte pentru a-ți maximiza experiența de joc în Verde Casino. Fiind o oază a bonusurilor, această platformă își păstrează promisiunea de a răsplăti jucătorii într-un mod generos și captivant.
Verde casino bonus pentru clienții noi
Verde Casino își răsplătește generos jucătorii noi printr-un program de bonusuri la depozit captivant. În cadrul ofertei de bun venit, aceștia pot beneficia de următoarele avantaje:
| Bonus la prima depunere: | Primiți un bonus de 120%, cu o valoare maximă de 1200 lei. Bonusul este însoțit de 50 de rotiri gratuite la slotul Big Bass Bonanza. |
| Bonus la a doua depunere: | Beneficiați de un bonus de 150%, cu o valoare maximă de 1200 lei. În plus, vă așteaptă 50 de rotiri gratuite la celebrul slot Book of Dead. |
| Bonus la a treia depunere: | Jucătorii primesc un bonus de 100%, până la suma de 1200 lei. În completare, se adaugă 50 de rotiri gratuite la aparatul Starburst. |
| Bonus la a patra depunere: | Un bonus atractiv de 150%, cu o valoare maximă de 1200 lei. Alături de acesta, sunt oferite 70 de rotiri gratuite la slotul Book of Fallen. |
Aceste bonusuri substanțiale asigură un start de excepție în experiența de joc la Verde Casino, oferind nu doar bani suplimentari, ci și șanse gratuite la câștiguri consistente prin rotirile gratuite la sloturi populare.
Bonus săptămânal
La Verde Casino, jucătorii fideli au oportunitatea de a profita săptămânal de un bonus generos în valoare de până la 4000 Ron și 100 de rotiri gratuite. Pentru a beneficia de această ofertă tentantă, trebuie să atingi nivelul Golden Overlord în cadrul programului de loialitate al cazinoului.
Bonus cashback
La Verde Casino, avem grijă și de pierderile tale! Prin programul nostru de cashback, poți recupera până la 12% din sumele pierdute în fiecare săptămână, cu o limită maximă generoasă de 8000 Ron. Iată mai multe detalii despre condițiile și cerințele acestui bonus cashback:
Condiții pentru Bonus Cashback:
- Înregistrare: Asigură-te că ești înregistrat la Verde Casino și ai un cont activ.
- Program de Loialitate: Avansează în cadrul programului de loialitate al cazinoului și atinge nivelul Diamond Overlord.
Programul de loialitate
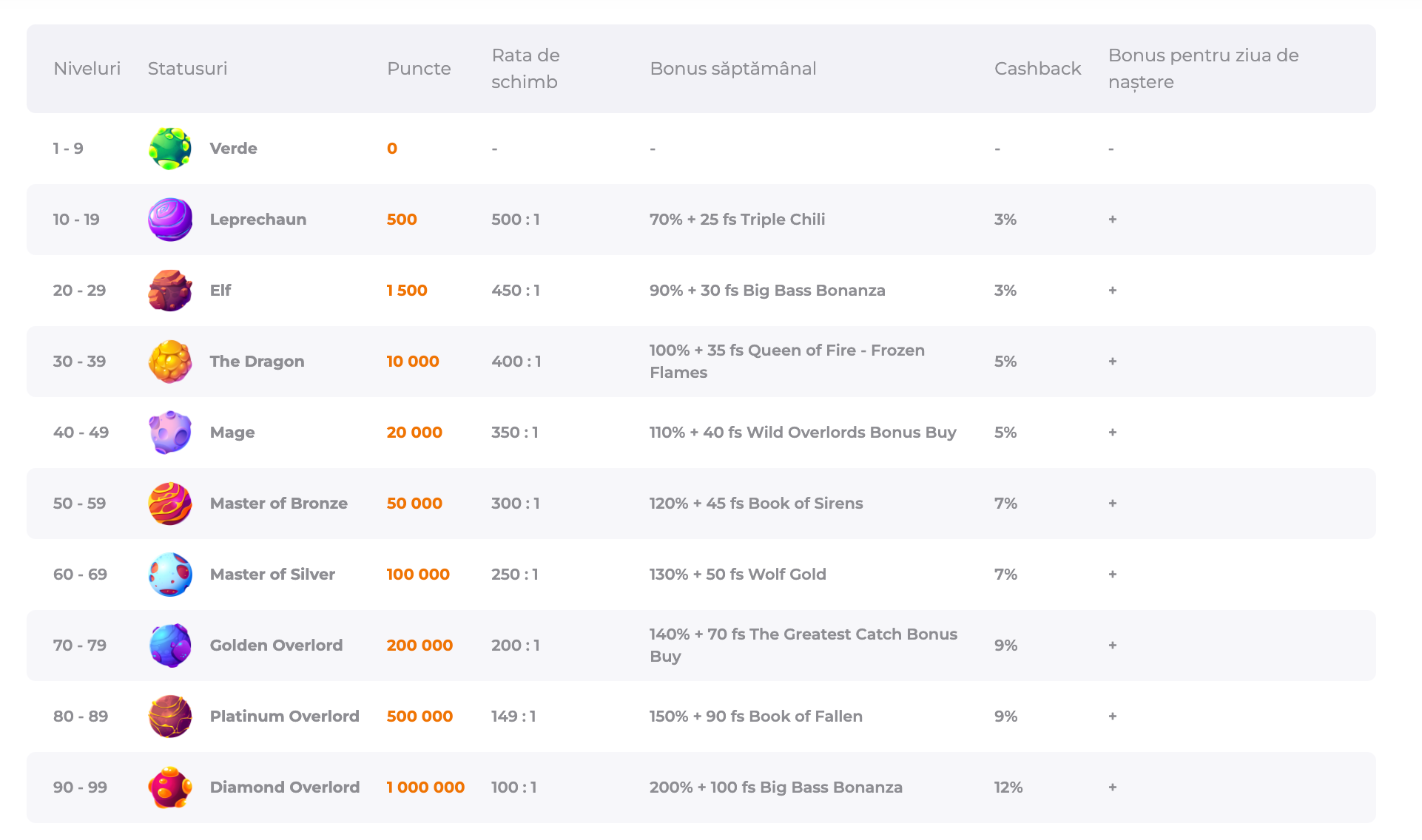
În calitate de membru Verde Casino, vei experimenta nu doar distracție intensă, ci și beneficiile programului nostru de loialitate. Odată ce te-ai înregistrat și ai efectuat prima depunere, obții nu doar un avantaj imediat sub formă de bonus de bun venit, ci și acces la programul nostru de loialitate, care aduce recompense pe măsură ce avansezi în nivele. Iată cum funcționează:
Acumularea Punctelor de Fidelitate:
- Fiecare 40 Ron rulați la jocurile cazinoului echivalează cu 1 punct de fidelitate.
- Punctele acumulate deschid ușa către numeroase beneficii și recompense.
Exemple de Nivele în Programul de Loialitate:
- Niveluri Inițiale: Verde, Leprechaun, Elf.
- Niveluri Intermediare: The Dragon, Mage, Master of Bronze, Master of Silver.
- Niveluri Superioare: Golden Overlord, Platinum Overlord, Diamond Overlord.
Recompense și Bonusuri:
- Bonus la Reîncărcarea Contului: Între 10% și 90% din sumă.
- Bonus Cashback: Între 3% și 12% din sumele pierdute săptămânal.
- Rotiri Gratuite: Între 25 și 75 runde gratuite.
- Bonus Aniversar: O surpriză specială la aniversarea ta.
Premii în Bani Cash Săptămânale:
- Până la 2000 Ron pentru nivelele Leprechaun și Elf.
- Până la 2800 Ron pentru nivelele The Dragon, Mage, Master of Bronze și Master of Silver.
- Până la 4000 Ron pentru nivelele Golden, Platinum și Diamond Overlord.
Prin acest program de loialitate, Verde Casino îți oferă o gamă variată de recompense, de la bani cash și rotiri gratuite la bonusuri aniversare. Cu fiecare punct de fidelitate acumulat, te apropie mai mult de statuturi superioare și de beneficiile lor atractive. Intră în aventura ta la Verde Casino și bucură-te de recompensele pe care le aduce programul nostru de loialitate!
Verde la super jocuri de top cu platforma online Verde Casino
Verde Casino oferă o gamă variată de opțiuni de joc, cu peste 3000 de sloturi și peste 100 de jocuri de masă, fiecare certificat pentru RNG și fair play. Iată o privire mai detaliată la secțiunile principale de jocuri și exemple de titluri atrăgătoare:
| Sloturi | Descoperiți o selecție vastă de păcănele încercate și testate. Recomandări: Big Bass Bonanza și Sweet Bonanza de la Pragmatic Play. |
| Jocuri de Masă | Găsiți toate opțiunile preferate, inclusiv zaruri, ruletă, plinko, keno, poker, ruletă și blackjack clasic. Încercați-vă norocul la Triple Edge Poker sau Andar Bahar de la Swintt. |
| Casino Live | Duceți-vă experiența la un nivel superior cu dealeri live. Mese live de la Pragmatic Play, Absolute Live Gaming, Swintt și Evolution Gaming. Experimentați și mesele live show TV, cum ar fi Mega Ball, Monopoly Live sau Crazy Coin Flip. |
Indiferent de preferințele dvs., Verde Casino vă asigură că veți găsi o varietate de jocuri captivante, de la cele clasice la cele inovatoare, create de furnizori de top în industrie. Jucați în siguranță și cu încredere, având în vedere certificările pentru fair play ale fiecărui titlu disponibil.
Sloturi online
La Verde Casino, sloturile sunt vedetele principale, oferindu-vă o experiență autentică de cazino online. Portofoliul bogat include o varietate de opțiuni, de la sloturi clasice la păcănele populare, video sloturi cu linii de plată multiple, până la pacanele Megaways, toate cu tematici captivante și distractive. Printre furnizorii noștri de top se numără Evoplay, Spinomenal, Gamomat, Endorphina, Betsoft și mulți alții, recunoscuți pentru jocurile lor apreciate la nivel global. Aceasta este doar începutul, deoarece ne propunem să adăugăm continuu noi creatori de păcănele pentru a vă oferi mereu ceva nou și captivant în experiența dvs. de joc. La Verde Casino, jocurile de slot sunt cu adevărat acasă!
Ruletă
La Verde Casino, ruleta este o experiență captivantă, indiferent dacă alegi varianta standard sau cea live. Te așteaptă o selecție variată de opțiuni, inclusiv ruleta europeană și americană, în diferite versiuni pentru a satisface orice preferință. În plus față de variantele clasice, poți încerca și variante mai exclusive, precum VIP Roulette, Zoom Roulette, First Person Lightning Roulette sau XXXtreme Lightning Roulette. Cu furnizori de top la bord, experiența ta la ruletă va fi una autentică și plină de emoție, doar la Verde Casino.
Poker
Dacă ești un pasionat al jocurilor de minte, Verde Casino îți oferă o experiență bogată de poker în diverse variante. Aici, poți explora jocuri precum Oasis Poker, Pai Gow Poker, Triple Edge Poker și Caribbean Poker, toate concepute pentru a-ți stimula abilitățile strategice și pentru a-ți oferi momente de divertisment captivante. La Verde Casino, pokerul devine o adevărată provocare intelectuală și o modalitate excelentă de a-ți testa norocul și priceperea într-un mediu online captivant.
Jocuri Instant

Dacă nu-ți poți stăpâni curiozitatea și vrei să descoperi imediat rezultatul, atunci jocurile instant de la Verde Casino sunt alegerea potrivită pentru tine. Descoperă dacă ai câștigat într-o clipă cu opțiuni precum Aviator, Mines, Keno sau Plinko, sau explorează experiența de cazino online cu titluri captivante precum Lucky Score, Champions Pick sau Scratchy Bit. La Verde Casino, adrenalina câștigului este la doar un clic distanță!
Jocuri cu dealeri reali live
Descoperă atmosfera autentică a cazinoului la Verde Casino, unde te așteaptă o secțiune de jocuri cu dealeri reali în parteneriat cu Evolution Gaming și Pragmatic Play, lideri mondiali în domeniul live casino. Experimentează senzația de joc la mesele de ruletă, blackjack, și baccarat, bucurându-te de o varietate de opțiuni captivante. La Verde Casino, oferim o experiență autentică de cazino online, adusă în confortul casei tale.
Jocuri de masă și de cărți
Descoperă diversitatea și excitarea în secțiunea noastră de jocuri de masă și de cărți la Verde Casino online. Îți punem la dispoziție o selecție generoasă care acoperă atât variantele clasice, cât și cele mai puțin cunoscute. Bucură-te de jocuri populare precum blackjack, poker, și ruletă, dar și de opțiuni mai inedite, inclusiv Casino Barbut, Casino Patience, Red Dog, Dice, Hi-Lo, și Keno. La Verde Casino, avem o alegere pentru fiecare pasionat de jocuri de masă și de cărți.
Verde Casino Mobile
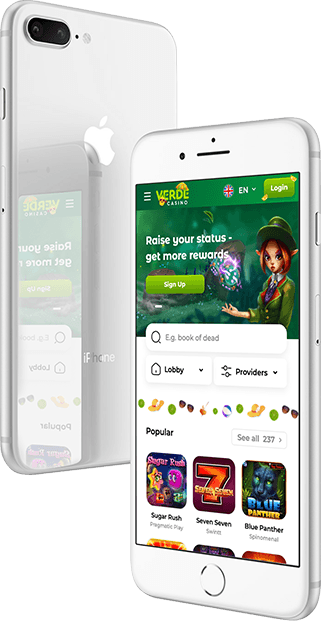
Jucătorii care preferă experiența mobilă la Verde Casino beneficiază de un confort deosebit, având opțiunea de a plasa pariuri oricând și oriunde. Platforma mobilă poate fi accesată fie prin intermediul site-ului, fie prin intermediul unei aplicații adaptabile. Pentru cei interesați de Verde Casino download, aplicația este disponibilă pe pagina oficială a cazinoului. Aceasta oferă economie de trafic și încărcare rapidă a interfeței. Procesul de instalare pentru Verde Casino apk implică găsirea secțiunii cu kitul de distribuție pe site, descărcarea apk-ului, permiterea instalării din surse necunoscute și finalizarea procesului de instalare. Utilizatorii Android pot instala Verde Casino apk în doar 2-3 minute, ocupând un spațiu redus pe dispozitiv. În prezent, Verde Casino iOS nu este disponibil, însă utilizatorii de dispozitive iOS pot accesa versiunea mobilă, echivalentă în funcționalitate și varietate de jocuri cu site-ul oficial.
Verde Casino sistemele de plată

Cu Verde Casino, procesul de înregistrare se desfășoară într-un ritm alert, asigurându-ți accesul rapid la diversitatea de jocuri și servicii. La fel de important precum viteza înregistrării sunt și metodele de plată eficiente și securizate oferite pentru a facilita experiența ta pe platforma noastră. Iată o listă a metodelor de plată disponibile:
- MiFinity Voucher
- Transfer rapid
- Skrill
- Neteller
- Neosurf
- Bitcoin
- Tether
- Biance Pay
- Astro Pay
Procesul de depunere este simplu: autentifică-te în contul tău Verde Casino, alege o modalitate de plată (online banking, portofel electronic sau criptomonede), selectează suma dorită, confirmă tranzacția, iar banii vor fi disponibili în contul tău în timp util.
Pentru retragerea câștigurilor, este necesar să confirmi adresa de email, să furnizezi datele complete de identitate și să treci prin procesul KYC (Know Your Customer) oferind dovezi ale identității reale, adresei reale și a unei metode de plată reale înregistrate pe numele tău. Ulterior, retragerea banilor se realizează în mod simplu și eficient, folosind aceeași metodă ca la depunere. La Verde Casino, ne străduim să oferim o experiență fluidă și rapidă, astfel încât membrii noștri să se bucure la maximum de jocurile noastre.
| Metodă de depunere | Timp procesare | Depunere minimă |
| Visa/MasterCard | instant | 50 RON / 10 USD |
| Skrill | instant | 50 RON / 10 USD |
| Neteller | instant | 50 RON / 10 USD |
| Paysafecard | instant | 50 RON / 10 USD |
| ecoPayz | instant | 50 RON / 10 USD |
| Metodă de retragere | Timp procesare | Retragere minimă |
| Visa/MasterCard | 5 minute și 5 zile | 100 RON / 20 USD |
| Skrill | 5 minute și 5 zile | 100 RON / 20 USD |
| Neteller | 5 minute și 5 zile | 100 RON / 20 USD |
| Paysafecard | 5 minute și 5 zile | 100 RON / 20 USD |
| ecoPayz | 5 minute și 5 zile | 100 RON / 20 USD |
Serviciul de asistență online Verde Casino
Suntem dedicati oferirii unui suport complet și prietenos la Verde Casino, chiar dacă momentan asistența este disponibilă predominant în limba engleză. Cu toate acestea, ne-am asigurat că serviciul nostru de chat live este accesibil și ușor de utilizat pentru toți jucătorii noștri vorbitori de limba română.
Chatul live este activ 24/7 și poți accesa acest serviciu de pe orice dispozitiv conectat la internet, indiferent dacă folosești un iPad, iPhone, Android, Windows Mobile, PC sau Mac. Accesează canalul de chat apăsând butonul portocaliu din colțul dreapta jos al paginii. Alege una dintre întrebările cu caracter general dacă găsești răspunsul în acea secțiune sau apasă pe butonul “Open Chat” pentru a intra în direct cu un operator pregătit să răspundă întrebărilor tale în timp real. La Verde Casino, suntem aici pentru tine, indiferent de întrebările sau preocupările pe care le ai.
Verde Casino Security
La Verde Casino, ne-am angajat ferm să asigurăm un mediu de joc sigur și de încredere pentru toți jucătorii noștri. În calitate de jucător, transparența și securitatea informațiilor tale sunt prioritare pentru noi.
Licența noastră provine de la guvernul Curacao și este verificabilă prin intermediul unui validator oficial. Această licență reprezintă o măsură esențială pentru a asigura că operăm în conformitate cu standardele industriei și că oferim o experiență de joc corectă și legală.
Deși certificatul eCOGRA nu este specificat, ne bazăm pe criptarea datelor cu o tehnologie avansată de 128 de biți pentru a proteja informațiile personale și financiare ale jucătorilor noștri. Aceasta adaugă un strat suplimentar de securitate, asigurând că toate datele sunt protejate împotriva accesului neautorizat.
Caracteristici de joc responsabil
La Verde Casino, suntem profund angajați în promovarea jocului responsabil și în asigurarea unei experiențe de divertisment sănătoase pentru toți jucătorii noștri. Deși este adevărat că nu avem o secțiune dedicată exclusiv pariurilor responsabile, dorim să subliniem importanța acestui aspect și să oferim suportul necesar jucătorilor noștri.
Încurajăm jucătorii să-și mențină controlul asupra experienței lor de joc prin stabilirea unor limite zilnice de pariuri. Segmentarea sesiunilor lungi de joc în intervale mai scurte poate contribui la menținerea unei abordări echilibrate și responsabile față de jocurile de noroc online.
În plus, recunoaștem importanța informațiilor despre ajutorul disponibil pentru cei care resimt primele semne ale dependenței de jocuri de noroc. Încurajăm jucătorii să caute resurse externe specializate care oferă suport și consiliere în această privință.
Opinii despre Verde Casino
Elena33:
- “Sunt jucătoare de cazinouri online de ceva timp, iar Verde Casino mi-a atras atenția cu adevărat. Ce îmi place cel mai mult este varietatea lor impresionantă de jocuri de slot și mese de joc. În plus, procesul de înregistrare a fost atât de rapid și facil, iar asistența clienți a fost de fiecare dată la înălțime. Recomand cu încredere!”
Alex82Poker:
- “Pasionat de poker, am descoperit la Verde Casino o gamă vastă de variante, de la clasicele jocuri de cărți până la versiunile mai inedite. Interfața live pentru poker este extraordinară, iar dealerii sunt profesioniști. Sistemul lor de loialitate m-a fidelizat și mai mult. Un loc minunat pentru fanii pokerului!”
Adrian_SlotMaster:
- “Sloturile sunt pasiunea mea, iar Verde Casino a devenit rapid destinația mea preferată. Se remarcă prin selecția lor diversificată și colaborarea cu furnizori de top. Programele lor de bonus sunt foarte generoase, iar plățile sunt rapide și eficiente. Recomand tuturor iubitorilor de sloturi!”
Roxana_RuletaFan:
- “Am găsit o adevărată oază a jocurilor de ruletă la Verde Casino. Variațiile lor m-au captivat, iar secțiunea live este pur și simplu fascinantă. Interacțiunea cu dealerii live adaugă o dimensiune nouă experienței mele de joc. Îmi place că site-ul este ușor de navigat și prietenos pentru utilizatori.”
GigiBetMaster:
- “Dacă ești pasionat de pariuri sportive, Verde Casino este locul potrivit. Gama lor de evenimente sportive este vastă, iar opțiunile de pariere sunt diversificate. Cotele sunt competitive, iar secțiunea de statistici m-a ajutat mult în luarea deciziilor. Un mediu prielnic pentru cei pasionați de sport și pariuri.”
BiancaCasinoExplorer:
- “Verde Casino m-a impresionat cu adevărat cu interfața lor prietenoasă și responsivă pentru mobil. Posibilitatea de a juca oricând și oriunde este un avantaj major. Am avut experiențe plăcute cu metodele lor de plată rapide și sigure. Bonusurile și promoțiile constante sunt un mare plus. Recomand tuturor căutătorilor de distracție online!”
Concluzie
În încheiere, Verde Casino se remarcă ca o destinație vibrantă și captivantă pentru iubitorii de jocuri de noroc online. Cu o gamă impresionantă de sloturi, jocuri de masă, opțiuni live și pariuri sportive, cazinoul oferă o experiență diversificată și plină de adrenalină. Programele lor generoase de bonusuri și promoțiile constante adaugă un element suplimentar de entuziasm pentru jucători, iar sistemul lor de loialitate recompensează fidelitatea într-un mod atractiv. Interfața intuitivă, adaptată pentru mobil, și varietatea de opțiuni de plată eficiente completează oferta, oferind o experiență completă și comodă. Cu serviciul lor de asistență disponibil 24/7 și accentul pe securitatea jocului, casino Verde se impune ca o alegere solidă pentru cei în căutare de divertisment online de calitate.
Alternativ, Conticazino este un cazinou online popular în România.
FAQ
🌿 Ce sisteme de plată sunt prezentate în cazinou?
Verde Casino oferă o varietate de metode de plată, inclusiv MiFinity Voucher, Transfer rapid, Skrill, Neteller, Neosurf, Bitcoin, Tether, Biance Pay, și Astro Pay.
🌿 Care sunt limitele pentru tranzacții?
Limitele pentru tranzacții pot varia în funcție de metoda de plată aleasă și statutul de membru. Aceste informații detaliate sunt disponibile în secțiunea de depunere și retragere a site-ului.
🌿 Ce jocuri sunt prezentate în cazinou?
Verde Casino dispune de o gamă diversificată de jocuri, inclusiv peste 3000 de sloturi, 100+ jocuri de masă, opțiuni live cu dealeri reali, jocuri instant, poker, ruletă, și multe altele, oferite de furnizori precum Evoplay, Spinomenal, Gamomat, și alții.
🌿 Este sigur să pariezi la un cazinou?
Da, Verde Casino este considerat sigur, având licență emisă de guvernul Curacao și criptare a datelor pe 128 de biți. Se acordă o atenție specială securității și confidențialității jucătorilor.